| Механизмы интеграции. |
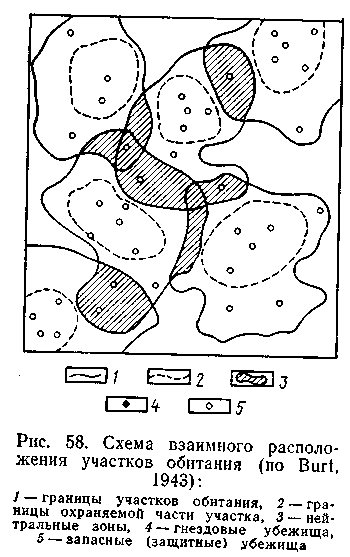
Различные способы индивидуализации территории пространственно разграничивают особей в составе популяции. Однако этот процесс не может продолжаться бесконечно: при слишком большой дисперсности теряется возможность поддержания устойчивых внутрипопуляционных контактов, на которых основываются целостность популяции и ее функции. Поэтому в поддержании пространственной структуры популяций механизмы, определяющие возможность контактов между особями, их интеграцию в единую систему, имеют не меньшее биологическое значение. Эти механизмы направлены на ограничение дисперсности особей в пространстве, «уравновешивая» действие механизмов индивидуализации территории. В основе механизмов интеграции лежит наследственный стереотип поведения, связанный со стремлением войти в контакт с особями своего вида. Информация о присутствии особей в виде различных сигналов, следов жизнедеятельности, характерных изменений рельефа и растительности создает в местах постоянного обитания животных «биологическое сигнальное поле» (Наумов, 1973, 1975), которое служит мощным стимулом при выборе мест поселения другими особями того же вида. Показано, например, что большие песчанки Rhombomys opimls охотнее селятся даже на пустующих колониях того же вида, нежели в аналогичной местности по соседству; подобное явление известно для многих других животных. В опытах с монгольскими песчанками показано, что подчиненные животные не только не избегают запаха доминанта, но, напротив, предпочитают его (Tang, 1978). У ряда видов водных беспозвоночных (морские уточки Balanus sp. и др.) подвижные личинки оседают и продолжают развитие преимущественно в тех местах, где уже прикрепились другие особи; так, рассеянные в толще воды личинки формируют колонии прикрепленных взрослых форм. Икрометание многих рыб определяется местом откладки первых порций икры. Это явление специально исследовалось в экспериментах с Cichlasoma nigrifasciatum. В качестве субстрата для прикрепления икры рыбам предлагались три пластинки, расположенные в различных плоскостях. При откладке первых порций икры статистически значимого предпочитания какой-либо из пластинок не обнаруживалось. Если же на одной из них (независимо от ее положения) предварительно располагали кладку, то рыбы с высокой степенью достоверности откладывали икру именно на эту пластинку (Weber, Weber, 1971). В полевых экспериментах с лесной лягушкой Rana sylvatica кладку икры перенесли в пруд, в котором икрометание еще не началось. Через несколько дней все отложенные в этом пруду кладки оказались в непосредственной близости от интродуцированной кладки (Howard, 1980). Известно, что головастики бесхвостых амфибий образуют скопления. В экспериментах с Btifo americanus было показано, что они формируются не случайно: при запуске в общий аквариум головастиков разных кладок они вскоре группировались по «родственному» принципу (скопления из головастиков одной кладки), ориентируясь, скорее всего, на химические сигналы (Waldham, Adler, 1979). В лабораторных условиях установлено, что ящерицы анолисы Anolis auratus стремятся присоединиться к особям своего вида Ориентация в этом случае идет, видимо, на основе зрительной рецепции, поскольку при отсутствии особей своего вида ящерицу присоединялись к очень похожим внешне представителям близкого вида A. tropidogaster (Kiestor 1979). Все эти наблюдения и эксперименты позволяют заключить, что активное стремление к контактам с особями своего вида действует как механизм пространственной интеграции, объединения популяции. В сложившихся популяциях активный поиск информации и контактов составляет одну из характерных сторон исследовательского поведения животных. Формы исследовательской активности весьма разнообразны. Так, при передвижениях рыжих полевок по участку обитания, помимо коротких выходов з определенные места кормежки, удается различить особый «патрулирующий» тип передвижений: после выхода из норы зверек в быстром темпе обегает весь участок (особенно его периферические части), задерживаясь на короткое время в некоторых точках, помеченных мочой (Карулин и др., 1973). Сходный тип поведения наблюдался у малых сусликов и ряда других грызунов. Регулярные облеты границ участка характерны для многих птиц, аналогичное явление встречается и у других животных. Биологическое значение «патрулирования» участка заключается в получении информации о соседних особях и оставлении своих меток. Роль информационных контактов настолько велика, что утрата их может быть причиной разрушения сложившейся структуры. Так, при экспериментальном истреблении монгольских песчанок одна группировка этих животных была намеренно сохранена. Несмотря на то, что связи особей, составляющих группировку, не были нарушены, она вскоре распалась - видимо, в силу утраты привычной информации со стороны соседних групп. Увеличение подвижности в популяциях грызунов при депрессии численности - явление известное; оно связано с поиском контактов, в процессе которого образуется новая форма пространственного распределения. В результате тяготения к устойчивым контактам полное разобщение участков обитания отдельных особей скорее исключение, чем правило. У большинства видов участки соседних особей в той или иной степени перекрываются. При этом зона перекрытия в равной степени относится к участкам каждой из соседних особей, маркируется ими. Это возможно потому, что в большинстве случаев активно охраняемая территория охватывает не всю площадь участка, а только часть ее, тогда как различные формы использования (питание, маркировка, укрытие от хищников) распространяются на весь участок (рис. 58).
Зона перекрывания двух или нескольких смежных участков представляет собой своеобразную «зону контакта», на которой животные-соседи не проявляют агрессивности по отношению друг к другу. Так, наблюдения за малыми сусликами показывают, что на общей части территории зверьки-соседи мирно кормятся рядом, при опасности прячутся в соседние защитные норы, совместно нападают на «чужаков»; агрессивная реакция возникает лишь тогда, когда один из сусликов проникает на ту часть территории, которую сосед использует один. Аналогичные наблюдения имеются и по отношению к другим видам. У волков, например, «буферные» зоны посещаются обеими соседними стаями, но лишь на короткое время. При обилии добычи ни одна стая в этих зонах не охотится; это послужило основанием для гипотезы, согласно которой «буферные» зоны функционируют как место беспрепятственного возобновления популяций жертв (Mech, 1977).
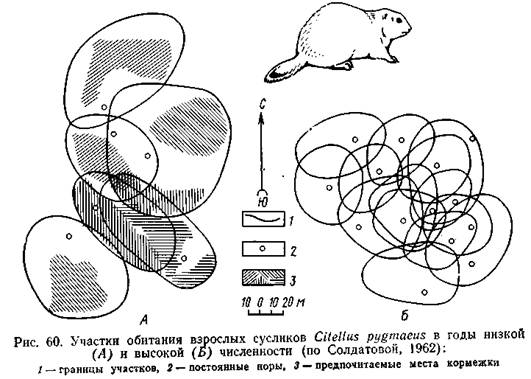
Немалое значение для увеличения интегрированности особей в популяции имеет и дифференцированный характер проявления территориального поведения. Как правило, участок активно защищается от «равного» партнера, потенциального конкурента. У ряда видов рыб, рептилий и некоторых других животных территориальная агрессия возникает между одноразмерными особями и не выражена по отношению к более мелким. По-разному реагирует особь-хозяин на самцов и самок, проявляя терпимость к особям противоположного пола, по крайней мере в период репродуктивной активности. Во многих случаях регистрируются терпимые взаимоотношения между взрослыми оседлыми особями и проникающим на их участки молодняком: это показано для ряда видов рептилий, птиц и млекопитающих. В отношении млекопитающих экспериментально установлено, что возраст животных различается на основе обонятельной рецепции. В частности, у мышевидных грызунов состав секрета кожных запаховых желез меняется с возрастом: молодые животные обоих полов издают запах, подобный запаху взрослых самок; у достигших половозрелости самцов запах усложняется и приобретает специфический характер. Видимо, запаховым сходством с самками и объясняется терпимое отношение взрослых самцов к неполовозрелым животным (Stoddart, 1974). Несовпадение площади участков обитания с их активно охраняемой частью и вытекающая отсюда возможность перекрывания участков предоставляют популяции возможность «маневра» в использовании пространства. В частности, при нарастании численности и плотности населения участки обитания отдельных особей, как правило, уменьшаются в размерах (рис. 60).
Это возможно потому, что обычно величина участков обеспечивает их обитателям избыточное количество кормовых и иных ресурсов. Такое явление описано для многих видов; удалось установить, что при общем сокращении размеров участка особенно выражено уменьшение площади его охраняемой части, что открывает возможность прогрессивного увеличения степени перекрывания участков. Другой результат активного поиска контактов и перекрывания участков обитания заключается в том, что на этой основе формируются внутрипопуляционные группировки, в которых особи находятся в устойчивых закономерных взаимоотношениях. Внутрипопуляционные группировки служат структурной основой стабильности и «управляемости» популяции как биологической системы. Многочисленные наблюдения за различными видами животных из разных таксонов показывают, что даже в относительно однородных условиях местности внутри популяции формируются группировки особей, более тесно связанных друг с другом, нежели с другими аналогичными группировками. Значение таких агрегаций впервые отметил Н. П. Наумов, назвавший их «парцеллами» или «парцеллярными группировками». По Н. П. Наумову, парцеллярные (мы будем их называть более нейтральным термином «внутрипопуляционные») группировки представляют собой совокупность особей и семей, живущих в ближайшем соседстве и непосредственно связанных друг с другом теми или иными формами сигнализации. Они возникают активно, и их биологическая роль заключается в различных формах взаимопомощи (Наумов, 1967). Дальнейшие исследования показали, что роль внутрипопуляционных группировок еще более значительна: они представляют собой элементарную структурную единицу популяции, и именно на их основе обеспечивается адаптивный ответ популяции в целом на изменение вне- и внутрипопуляционных условий (Шилов, 1977). Несмотря на «индивидуальную» неустойчивость таких группировок (они могут распадаться и формироваться вновь; состав индивидов в каждой группировке может все время меняться), именно их существование обеспечивает устойчивость популяции в целом. Основой для этого служит система закономерных взаимоотношений особей в группировках, которая обозначается как «этологическая-» (функциональная) структура популяции. |