| Механизмы «индивидуализации» территории. |
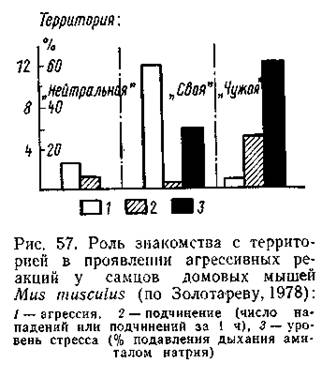
Биологические преимущества обитания на постоянном участке могут быть реализованы только при условии «индивидуализации» территории (Солдатова, 1972), использования ее только обитающими здесь особью или семьей. Наиболее прямая форма территориальных взаимоотношений представлена генетически детерминированным стереотипом агрессивного поведения, направленного на особей своего вида, проникших на территорию участка. ерриториальная агрессия в той или иной форме свойственна всем видам, имеющим участки обитания, в том числе и беспозвоночным (некоторые насекомые, ракообразные и др.). Биологически важно, что при территориальных конфликтах победителем (по крайней мере, у позвоночных) в подавляющем большинстве случаев оказывается особь, на участке которой произошла встреча (особь-резидент, «хозяин» данного участка). Подробные наблюдения за индивидуально помеченными малыми сусликами в естественных популяциях показали, что суслик, кормящийся в пределах своего участка, немедленно бросается на другого зверька (даже визуально знакомого соседа), как только тот пересечет невидимую, но строго соблюдаемую границу индивидуального участка. При этом пришелец, как правило, тотчас обращается в бегство, преследуемый «хозяином» до его границ, после чего погоня прекращается. В отдельных случаях, когда «хозяин», увлекшись погоней, пересекает эту границу, положение меняется: убегающий зверек, оказавшись на своей территории, переходит в нападение и изгоняет своего преследователя (Солдато-ва, 1962, 1966, 1967). Очень демонстративны опыты, проведенные в естественных условиях с мухоловками-пеструшками Muscicapa hypoleuca. Дуплянку с гнездом этих птиц постепенно перемещали, пока она не оказывалась на территории гнездового участка соседней пары. Хозяева участка немедленно начинали атаки и оказывались победителями в возникающих схватках. После восстановления исходного положения передвигали гнездовье победителей на территорию побежденной пары. При этом активной стороной были птицы, территория которых подверглась вторжению, они и одерживали победу. Если обеих дуплянок сближали на «нейтральной» (находящейся вне пределов обоих участков) территории, агрессивных взаимоотношений менаду птицами не отмечалось (Благосклонов, 1962, 1964). Аналогичные результаты были получены и в лабораторных экспериментах с различными видами рыб, землеройками Sorex araneus, белыми и домовыми мышами и др. Во всех случаях особи-хозяева проявляли отчетливую агрессивную реакцию, тогда как «пришельцы» либо не отвечали на нападение, либо их собственная агрессия была ослаблена. При этом разница в поведении зависела не от индивидуальных свойств данной особи, а именно от ее статуса: у одной и той же особи агрессия возрастала, если встречи происходили на освоенной ею территории, и ослаблялась на участке другого животного. Так, в опытах с зеленой саламандрой Aneides aeneus из 45 агрессивных встреч в 43 случаях победителями стали самцы, первыми освоившие экспериментальную камеру; «чужие» самцы проявляли агрессию лишь в 13 опытах, причем только в ответ на нападения резидента. Когда «хозяев» и «чужаков» поменяли ролями, агрессия и победы доминировали у новых хозяев (Сирр, 1980).
Все эти и многие другие данные подтверждают, что победа «хозяина» участка - закономерное явление, биологический смысл которого заключается в стабилизации сложившейся пространственной структуры популяции. Физиологические механизмы этого феномена изучены слабо. Можно полагать, что в основе предопределенного решения территориальных конфликтов лежат различия в мотивации поведения у особей, находящихся на знакомой территории, и зверьков, оказавшихся в непривычной обстановке. У первых доминирует врожденный стереотип территориальной агрессии, подкрепляемый всей системой знакомых ориентиров, среди которых появляется объект нападения. У «чужаков» на незнакомой территории развивается ориентировочная реакция - «специфическая целостная деятельность организма, в своем развитии затормаживающая все остальные деятельности» (Анохин, 1958), которая легко переходит в пассивно-оборонительную реакцию. Различная «ориентация» высшей нервной деятельности и решает исход конфликта в пользу «хозяина». Эта гипотеза подтверждается рядом полевых и лабораторных наблюдений. У сусликов, попавших на незнакомую территорию, резко меняется поведение: они передвигаются неуверенно, часто поднимаются «столбиком», проявляют все признаки ориентировочного рефлекса. Аналогичные формы поведения на незнакомой территории наблюдались и у многих других грызунов. Если создается обстановка, при которой ориентировочный рефлекс возникает и у «хозяина» участка, шансы партнеров выравниваются и агрессия снижается. В частности, в опытах с трехиглой колюшкой Gasterosteus aculeatus было установлено, что агрессивность территориального самца падает, если увеличивается число одновременно присутствующих противников (Van den Assen, 1970). Аналогичные наблюдения сделаны в опытах с землеройками, грызунами и некоторыми другими животными. Можно полагать, что одновременное присутствие нескольких соперников рассеивает внимание хозяина, вызывает у него ориентировочную реакцию и тем как бы уравнивает положение всех особей. Механизм подобного типа действует в течение относительно короткого времени. Важно, однако, что в природе исход встречи резидента с пришельцем решается именно в короткий срок, что вполне удовлетворительно объясняется рассмотренной гипотезой. Роль агрессии в поддержании пространственной структуры показана в полевых экспериментах с искусственным подавлением враждебного поведения. Серия таких опытов была проведена с монгольскими пищухами Ocholona pricei. Этим животным свойствен строго территориальный образ жизни, поддерживаемый отчетливыми агрессивными реакциями по отношению к соседним особям. В естественной популяции этих животных доминантам и субдоминантам вводили резерпин, подавляющий враждебное поведение. Это вызвало ослабление охраны участков, в результате чего некоторые особи были вытеснены за пределы семейной территории и началось хаотическое перемещение зверьков из одной норы в другую (Шилова, Туров, 1977). Аналогичные опыты с длиннохвостыми сусликами Ciiellus undulatus привели к сходным результатам. У оседлых зверьков снижалась агрессивность, а соответственно и способность к охране территории (Шилова и др., 1979). Монгольским песчанкам Meriones unguictdatus в летний период свойствен семейно-групповой образ жизни. Зверьки каждой семейной группы охраняют территорию участка от особей других семей. Введение подопытным зверькам галоперидола, снимающего агрессивность, значительно нарушало территориальное поведение. В результате произошло перераспределение территории между подопытными зверьками и положения их внутри семейных групп; в контроле подобных нарушений структуры не отмечалось (Громов, Попов, 1979). Все эти материалы подтверждают важную роль агонистического поведения в формировании и поддержании пространственной структуры популяций. Формы территориального поведения отнюдь не исчерпываются прямым нападением, схватками, погонями и т. д. Более того, такие «жесткие» формы встречаются не так уж часто. Прямая агрессия практически всегда сопровождается «ритуальными» формами поведения: позой угрозы, специальными звуковыми сигналами и пр. Значение угрозы имеет, например, определенный наклон туловища по отношению к горизонтали у ряда видов рыб, «приподнятые» позы у некоторых бесхвостых амфибий, «кивание», приподнимание и опускание передней части тела и расправление горловой складки у ящериц-игуанид, демонстрация ярких участков тела у некоторых других ящериц, многообразные угрожающие позы и движения у разных видов млекопитающих. В подавляющем большинстве случаев демонстрация угрозы воспринимается особью, попавшей на чужой участок, как сигнал к бегству - возможно потому, что этот сигнал действует на фоне развитого ориентировочного рефлекса. Во многих случаях территориальное поведение полностью исчерпывается демонстрациями, не переходящими в прямую агрессию. Так, установлено, что у коралловых рыб Pomacentris Jenkins сражения целиком носят ритуальный характер и почти полностью исключают физический контакт между партнерами. Такая «условная» форма агрессии широко распространена и в других группах животных. Еще более «мягкой» формой индивидуализации территории являются многообразные способы маркирования участков. У видов с хорошо развитым зрением нередко наблюдается визуальное мече-ние территории. Например, у видов коралловых рыб, занимающих небольшие участки с хорошими условиями видимости, яркая окраска с броским рисунком делает само присутствие рыбы достаточно ощутимым сигналом о занятости территории. В сочетании с демонстрационным поведением такая система маркирования вполне эффективна. Звуковая маркировка территории свойственна и другим животным. Так, взрослые самцы тюленя Уэделла Loptonychotes weddelii маркируют подводный участок вокруг щели, служащей им убежищем, с помощью особых трелей. Голосом метят групповой участок обезьяны-ревуны (p. Alouatta) и некоторые другие приматы. Звуковыми сигналами обозначают занятость участка американские бурундуковые белки. Широко используются акустические территориальные сигналы и у ряда беспозвоночных животных. У большинства млекопитающих наиболее важное значение в маркировке территории имеют запаховые метки, что соответствует ведущей роли обонятельной рецепции в жизни этих животных. «Носителем» запаха служат моча, экскременты или секреты специализированных желез. Тенреки Echinops telfairi метят предметы, нанося на них запах своего тела с помощью слюны: они попеременно трут лапой, намоченной слюной, свое тело (голову, бока) и участок маркировки (Eibl-Eibesfeldt, 1965). Запаховые метки оставляются по границам участка, а также на тропах, кормовых столиках, в районе убежищ и в других часто посещаемых местах. Установлено, что запаховая метка несет информацию не только о занятости территории, но и об индивидуальных особенностях данного животного (его пол, возраст, репродуктивная активность и т. д.). Выяснено, что специфический запах секретов различных желез иногда связан с деятельностью бактерий, способствующих анаэробному окислению секретов, в результате чего выделяются пахучие вещества (летучие жирные кислоты, амины и др.). Изменение секретов под действием бактерий может иметь определенные сигнальные функции (Albone et al., 1977). Наблюдается и «комплексная» система маркирования. Так, косули оставляют пахучие метки на стволах и ветвях, но, кроме того, метят свой участок голосом. Бурые медведи не только делают «задиры» и «закусы» на деревьях, но и наносят запаховые метки, потираясь о стволы холкой и головой. Бобры сочетают визуальное и химическое маркирование, устраивая на своих участках систему «запаховых холмиков». Эффективность различных способов маркировки участков определяется врожденным стереотипом негативной реакции на территориальный сигнал. Полевые наблюдения таких реакций, естественно, затруднены. Уже упоминалось о реакциях бегства в ответ на прямое или ритуализированное нападение у сусликов. Описано, как в колонии поморников Catharacta maccormiki новая пара отыскала место для гнездования и установила границы участка, ориентируясь на атаки соседей (Young, 1972). Были проведены полевые эксперименты с краснокрылыми трупиалами Agelaius phoeniceus, у которых территориальным сигналом служит демонстрация красных пятен на плечах («эполеты»). Выяснилось, что самцы, у которых «эполеты» были закрашены черным, показали ослабленную способность к защите территории; большинство из них были вытеснены со своих участков (Smith, 1972). Описана реакция избегания в ответ на визуальную метку (демонстрация контрастно окрашенного хвоста) у песчаной круглоголовки Phrynocephalus interscapularis. При полевых исследованиях биологии волков в США прямые наблюдения показали, что встреча с чужими запаховыми метками удерживает стаю от нарушения границы (Peters, Mech, 1975). В опытах с лабораторными мышами выявлено, что запах «чужих» самцов действует как стрессор: у подопытных особей масса надпочечников увеличивалась на 15-17%. У мышей с разрушенной обонятельной луковицей подобная реакция отсутствовала. Стрессовый эффект вызывается запахом выделяемого с мочой секрета придаточных желез половой системы, а также запахом секрета кожных желез подушечек лап (Ropartz, 1968). Не исключено, что стремление к снятию стресса - один из элементов поведенческой реакции на метку. Маркировка и территориальная агрессия тесно связаны с физиологией животных, в первую очередь с состоянием генеративной системы и соответствующим гормональным фоном в организме. Функционально активное состояние половой системы - мощный стимул к проявлению всего комплекса территориального поведения. Показано, например, что у колюшки Culaea inconstanus территориальное поведение стимулируется строительством гнезда. У многих видов рептилий и птиц поведение, связанное с защитой и маркировкой участка, наблюдается только в период размножения, и интенсивность его проявления зависит от стадии репродуктивного цикла. Такое же явление свойственно и некоторым млекопитающим (в частности, многим копытным); даже у видов, владеющих участками на протяжении всего года, интенсивность охраны и маркировки их вне периода размножения ослаблена. Показано, в частности, что специфические кожные подхвостовые железы у желтогорлых мышей Apodenius flavicollis развиты и функционируют только у готовых к размножению самцов. Регуляция маркировочного поведения с помощью половых гормонов биологически оправдана: именно в период размножения животные наиболее привязаны к территории, и индивидуализация ее особенно важна. У многих животных в ходе репродукции интенсивность маркировки снижается, в известной мере компенсируясь стереотипом привязанности каждого из соседей к своему участку, что снижает частоту нарушений их границ. Так, в частности, обстоит дело у многих видов птиц; то же, видимо, свойственно некоторым рыбам, амфибиям и другим животным, у которых участки образуются только на время размножения. Непосредственные стимулы, вызывающие маркировку участка, весьма разнообразны. В экспериментах установлено, что маркировка очень характерна при освоении нового помещения; в природе этому соответствует интенсивная маркировка при занятии участка. Очень широко распространена маркировка в ответ на метку чужой особи, что, вероятно, служит сигналом подтверждения границ участка в месте его соприкосновения с соседом. Существенно стимулирует мечение возбужденное состояние, вызванное усилением конкурентных отношений в популяции, появлением незнакомых особей, борьбой за самку и другими формами внутрипопуляцион-ных контактов. Анализируя изложенные способы индивидуализации территории, можно заметить, что они составляют последовательный ряд постепенно «смягчающихся» форм внутрипопуляционных отношений: от активной агрессии с нанесением противнику подчас тяжелых травм через ритуальные схватки с минимумом контакта к демонстрационному поведению без схваток и, наконец, к различным способам маркировки территории. Такая последовательность, разумеется, не отражает прямой путь эволюции территориального поведения, но тенденция к эволюционному закреплению более «мягких» его форм отчетливо проявляется во всех группах животных. Биологическое значение этого понятно: при всей значимости закономерного распределения территории для выживания отдельных особей и их потомства для популяции в целом важно, чтобы эта задача решалась с наименьшими потерями. |