| Газообмен в воздушной среде |
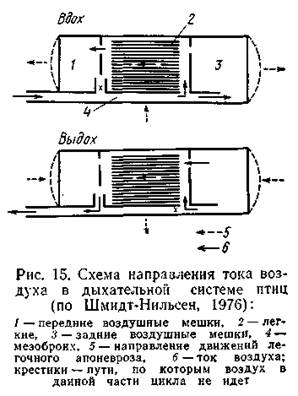
Кислород составляет 20,95% атмосферного воздуха. Соответственно велико и его парциальное давление: на уровне моря в условиях сухого воздуха эта величина равна 159,2 мм рт. ст. (21,2 кПа). Фактически парциальное давление О2 ниже, так как в воздухе всегда есть водяные пары, но все же оно остается достаточно высоким для эффективного газообмена на поверхности дыхательных органов. Фактором, лимитирующим газообмен в воздушной среде, оказывается сухость воздуха. Процесс непосредственного обмена га зов между кровью и внешней средой у наземных животных в принципе не отличается от газообмена у водных: в кровь поступает О2, предварительно растворенный в тонкой пленке влаги, покрываю щей поверхность дыхательного эпителия. Это обстоятельство определяет важную биологическую задачу поддержания дыхательной поверхности во влажном состоянии, что в условиях низкой и изменчивой влажности воздуха не всегда просто. Именно это условие и направило эволюцию строения и функции дыхательных органов наземных животных. Принципы воздушного дыхания. Морфологические принципы газообмена в воздушной среде строятся на том, что поверхность газообмена размещается внутри тела и не граничит непосредственно с окружающим воздухом. Большое число слизистых клеток поддерживает в дыхательной полости высокую влажность; дыхательные пути, связывающие органы дыхания с окружающей средой, также снабжены слизистым эпителием, что способствует увлажнению воздуха, поступающего в легкие. У позвоночных увлажнение вдыхаемого воздуха начинается уже в носовой полости, стенки которой выстланы слизистым эпителием, и поддерживается большим числом слизистых желез, расположенных в трахее и бронхах. У наземных беспозвоночных строение органов дыхания (их чаще всего тоже называют легкими) также подчинено этому принципу: дыхательная поверхность расположена внутри тела, отграничена от непосредственного соприкосновения с окружающим воздухом и соединена с ним узкими проходами. И у них поддерживается высокая влажность внутренних дыхательных поверхностей. У большинства наземных животных механизм дыхания представлен чередующимися фазами вдоха и выдоха. Такой тип дыхания, по-видимому, также обусловлен задачей сохранения высокой влажности в дыхательной системе: однонаправленный поток сухо го воздуха неминуемо приводил бы к подсыханию дыхательного эпителия и нарушению газообмена. Соотношение кожного и легочного дыхания у разных видов амфибий неодинаково и связано с их экологией: чем более сухие местообитания занимает тот или иной вид, тем большую относительную роль в газообмене играют легкие. Так, у прудовой лягушки Ranaesculentaпоступление О2 через кожу и легкие примерно одинаково (соответственно 51,2 и 48,8%), а СО2 выделяется преимущественно через кожу (86,2% от общего количества). У более наземной R. fuscaроль легких в потреблении О2 значительно выше (66,9%), тогда как СО2 и в этом случае в большем количестве (74,1%) выделяется через кожу. У хвостатых амфибий кожа представляет собой основной орган дыхания. Показано, что через кожу организм получает большую часть О2 и выделяет почти весь объем СО2. Легкие и ротоглоточная полость играют подчиненную роль: этим путем компенсируется недостаточное поступление О2в условиях, когда кожное дыхание затруднено (Северцов, Соколов, 1974). У видов, полностью перешедших к водному образу жизни, кожа выступает как основной орган водного дыхания. Легочное дыхание у амфибий более лабильно: сезонные изменения общего уровня газообмена у лягушек определяются главным образом динамикой легочного дыхания, тогда как кожное ос тается: практически неизменным в течение года; при этом в холодное время года преобладает кожное дыхание, в теплое -легочное. Неравноценна и роль двух систем дыхания в обмене газов: СО2 всегда энергичнее выводится через кожу, тогда как О2 следует этому правилу: лишь при низких температурах, а при высоких (выше 10°С) интенсивнее потребляется легочным путем. Дыхательные движения земноводных хорошо изучены на представителях бесхвостых. Они представляют собой довольно простую и не очень мощную систему всасывания воздуха в ротовую полость (где он перемешивается со смесью газов, выходящих из легких) и последующего проталкивания его в легкие. В этих движениях принимают участие мускулатура дна ротовой полости, связанная с подъязычным аппаратом, мышцы глотки, пережимающие ее в определенные моменты дыхательного цикла, и ноздри, которые могут открываться и закрываться. Вытеснение воздуха из легких происходит вследствие их эластичности. У рептилий строение легких в разных группах варьирует, демонстрируя усложнение внутренней структуры и, соответственно, увеличение суммарной дыхательной поверхности. Тип дыхательных движений у этих животных, в которых активно участвует грудная клетка, определяет более интенсивную, чем у амфибий, легочную вентиляцию. Лишь у некоторых форм, претерпевших вторичные изменения морфологии, эта схема несколько видоизменяется. У черепах, например, легкие вентилируются путем сокращения мышц, окружающих отверстия панциря, и «диафрагмы». У млекопитающих легкие имеют альвеолярное строение. Размеры альвеол невелики: например, у крысы их радиус в среднем составляет 20 мкм (для сравнения заметим, что у протоптеруса минимальный радиус альвеол - 50 мкм). Поэтому общая дыхательная поверхность оказывается очень большой, во много раз пре восходящей поверхность тела. Легочная масса состоит не только из альвеол, но и из подводящих структур (бронхи, бронхиолы). Объем этих структур, в которых остается часть отработанного воз духа, образует так называемое «мертвое пространство». Вдыхаемый воздух смешивается с воздухом мертвого пространства, по этому газовый состав воздуха в альвеолах отличается от газового состава атмосферного воздуха. Состав альвеолярного воздуха до вольно постоянен и у человека включает около 15% О2 и 5% СО2. Мощные дыхательные движения осуществляются не только хорошо развитой грудной клеткой, но и диафрагмой. Все это открывает возможность интенсивного газообмена и соответственно повышения уровня метаболизма. Гомойотермия млекопитающих в значительной степени определяется высоким уровнем окислительно-восстановительных процессов в организме. Дыхание у птиц. Строение дыхательной системы у птиц во многом отличается от дыхательной системы других наземных позвоночных. Легкие этих животных не имеют альвеолярного строения и отличаются малой растяжимостью. Их ткань представлена системой воздухоносных трубочек, открытых с обоих концов и потому допускающих возможность однонаправленного потока воздуха и соответственно непрерывного процесса газообмена. Дыхательная система, как и у других амниот, начинается трахеей и бронхами. Первичные бронхи входят в легкие, где дают ряд ответвлений (вторичные и третичные бронхи), и в конце концов открываются в воздушные мешки (тонкостенные полые образования, формирующиеся как выросты бронхов и располагающиеся между внутренними органами). Имеется 5 пар воздушных меш ков, которые функционально объединяются в две группы: передние (шейные, межключичные, переднегрудные) и задние (заднегрудные, брюшные). Воздушные мешки существенно увеличивают общий дыхательный объем и принимают активное участие в системе «воздушного насоса», прокачивающего воздух через легкие. Газообмен на внутренней поверхности воздушных мешков отсутствует. Дыхательный акт происходит с участием подвижных ребер и грудины и в принципе не отличается от грудного дыхания рептилий и млекопитающих. Прежнее представление о том, что в поле те дыхательные движения определяются взмахами крыльев, не соответствует действительности: точные эксперименты показывают, что в большинстве режимов полета синхронизация взмахов крыльев и дыхательных движений не соблюдается (По-видимому, лишь у голубя взмахи крыльев всегда синхронизированы с дыхательными движениями (Tomlinson, 1957); поскольку голубь - обычный объект физиологических исследований, это положение и было распространено на всех птиц. Как уже сказано, первичные бронхи (мезобронхи) образуют в легких многочисленные ответвления; от заднего их отдела отходят несколько вторичных бронхов, соединяющихся с передними воздушными мешками. В свою очередь, вторичные бронхи соединяются между собой многочисленными третичными бронхами (парабронхи). Каждый парабронх представляет собой тонкую трубочку (у цыпленка, например, диаметр ее составляет около 0,5 мм), от центрального канала которой отходят еще более тонкие канальцы - бронхиолы, делящиеся на воздухоносные капилляры диаметром 3-10 мкм. Бронхиолы и воздушные капилляры оплетены густой сетью кровеносных капилляров. Комплекс воздухоносных и кровеносных капилляров, образующих вокруг парабронхов сеть толщиной 50-200 мкм, представляет собой аппарат газообмена между воздухом, проходящим через легкие, и кровью. Подсчитано, что суммарная дыхательная поверхность легких курицы составляет 2,5 м2, т. е. примерно 1,1 м2/кг. Относительно меньшая, чем у млекопитающих, дыхательная поверхность определяется, видимо, большей эффективностью функции легких птиц (см. ниже). При вдохе наружный воздух поступает в задние воздушные мешки и частично в легкие; передние воздушные мешки на этой фазе дыхательного цикла заполняются воздухом из системы третичных бронхов, т. е. из собствен но легких. Вдыхаемый воздух в эту группу мешков не попадает (рис. 15). При выдохе мешки сжимаются и воздух из задних мешков выталкивается в легкие, т. е. в систему парабронхов, а из передних мешков - в трахею и через нее наружу (рис. 15, Б). На этой фазе дыхательного цикла воздух из задних мешков не поступает в первичные бронхи, а из передних - в легкие. Таким образом, через структуры легочной ткани воздух и при вдохе и при выдохе идет в одном направлении (от задних мешков к передним), что обеспечивает непрерывность процесса газообмена.
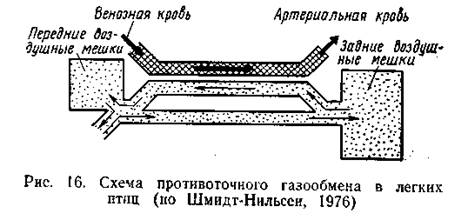
Респираторная система птиц лишена каких-либо клапанных систем, и направление воздушного потока по различным путям определяется относительной ориентацией входных отверстий различных воздухоносных путей, степенью их кривизны, а также изменением диаметра при вдохе и выдохе. Внутренние стенки парабронхов содержат гладкие мышечные волокна, сокращение которых способно регулировать их диаметр. В экспериментах с уткой по казано, что диаметр парабронхоз может изменяться в 2,3 раза, что соответствует почти тридцатикратной разнице в объеме воздуха, протекающего по сосудам в единицу времени. Предполагается, что автоматические изменения диаметра воздухоносных путей на разных фазах дыхательного цикла играют существенную роль в регуляции воздушного потока в респираторной системе птиц (Schmidt Nielsen, 1972). Важное преимущество проточной дыхательной системы птиц заключается в том, что при этом существенно повышается эффективность газообмена. В легких млекопитающих и других наземных позвоночных диффузия газов по градиенту парциального давления прекращается при выравнивании парциальных давлений газов в крови и в легочных альвеолах. В результате концентрация О2 и СО2 в артериальной крови всегда оказывается такой же, как в выдыхаемом воздухе. У птиц же напряжение СО2 в артериальной крови отличается большим постоянством и поддерживается на уровне более низком, чем содержание этого газа в выдыхаемом воздухе (в среднем около 30 мм рт. ст., т. е. 4 кПа). Обратные соотношения характерны для О2: птицы извлекают его из воздуха эффективнее, чем млекопитающие, и напряжение этого газа в артериальной крови выше, чем в выдыхаемом воздухе. Все это означает, что газообмен между воздухоносными и кровеносными капиллярами в легких птиц не ограничивается выравниванием напряжений этих газов в воздухе и в крови. К. Шмидт-Нильсен (Schmidt-Nielsen et al., 1969) высказал предположение о противоточной системе в легких птиц; упрощенная схема такой системы газообмена показала на рис. 16. Воздух, поступающий в легкие, из задних мешков имеет наиболее высокое парциальное давление О2. По мере прохождения по системе парабронхов воздух постепенно теряет О2 и получает СО2 из крови. При этом на всем протяжении этого пути воздух встречается с кровью, имеющей более низкое парциальное давление О2 и более высокое содержание СО2. Благодаря этому процесс встречной диффузии газов не прекращается и даже не замедляется, что и обеспечивает максимальную эффективность газообмена.
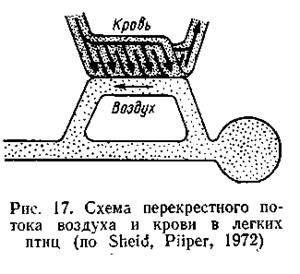
Более поздние исследования показали, что расположение воздухоносных и кровеносных капилляров в легких птиц несколько иное: каждый парабронх пересекается большим числом кровеносных капилляров, сеть которых оплетает его на всем протяжении (рис. 17). При такой системе перекрестного газообмена: градиент парциального давления газов также сохраняется по всей длине парабронха, что обеспечивает высокую эффективность извлечения О2 из воздуха (Sheid, Piiper, 1972).
Преимущества однонаправленного потока воздуха в легких птиц имеют особое значение при полете на большой высоте. Хорошая приспособленность птиц к низкому парциальному давлению О2былаэффектно продемонстрирована опытами в барокамере, в которой птиц (домовый воробей, волнистый попугайчик) и белых мышей «поднимали» до условий, соответствующих высоте 6100 м (атмосферное давление около 350 мм рт. ст., или 46,6 кПа, парциальное давление О2 73 мм, или 9,7кПа). В этих условиях при температуре 5°С через час после начала эксперимента воробьи увеличили уровень потребления О2 по сравнению с основным обменом в 2 раза и сохранили способность к короткому активному полету; лишь на «высоте» 7620 м они почти не мог ли летать, а на «высоте» 9140 м - даже стоять. Неакклиматизированные волнистые попугайчики так же сохранили способность к активному полету на «высоте» 6100 м. Мыши в этих опытах становились вялыми уже на «высоте» 3700 м, а при «подъеме» до 6100 м впадали в коматозное состояние (Tucker, 1968, 1972). Поскольку воробьи и мыши имеют сопоставимые размеры тела и почти одинаковый уровень основного обмена, а их кровь сходна по величинам кислородной емкости, неодинаковая реакция на низ кое давление может быть объяснена только разницей в принципах строения и функции легких у птиц и млекопитающих. Если бы газообмен у птиц, как и у млекопитающих, был ограничен выравниванием парциальных давлений газов воздуха и крови, то степень насыщения кислородом артериальной крови воробья на высоте 6100 м должна быть 24%; эти же данные, рассчитанные на основе принципа противоточного газообмена (Schmidt-Nielsen, 1972), да ют показатель насыщения 80%, что уже близко к нормальному. |