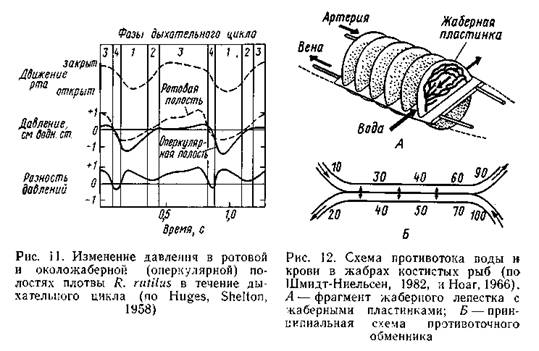
| Принцип водного дыхания |
Перечисленные особенности водной среды обусловливают соответствующее строение и функции дыхательной системы. У позвоночных животных, отличающихся относительно высоким уровнем обмена веществ, простой контакт дыхательной поверхности с водной средой (кожа, пассивно свешивающиеся в воду жабры), свойственный некоторым беспозвоночным, не в состоянии обеспечить необходимую скорость поступления кислорода в организм. У всех рыб жаберный аппарат устроен так, что вода активно прокачивается сквозь систему жаберных лепестков, через поверхность которых происходит газообмен; в менее эффективной форме это выражено у круглоротых. У костных рыб движения ротового и жаберного аппаратов сочетают нагнетательный (ротовая полость) и всасывающий (жаберная полость) принципы, что обеспечивает интенсивное продвижение воды сквозь жабры. У акуловых рыб благодаря отсутствию жаберной крышки этот механизм выражен слабее; при быстром движении, сопряженном с большими затратами энергии, для прокачивания воды через жабры используется само движение: рыба плывет с открытым ртом, и вода проталкивается через жабры тем интенсивнее, чем выше скорость движения. Такой же «пассивный» тип жаберной вентиляции отмечен и у быстро плавающих костистых рыб (например, у тун цов). Видимо, при большой скорости этот тип создания дыхательного потока воды оказывается более экономным. В опытах с полосатым окунем Moronesaxatilisи луфарем Pomatomussaltatrixвыяснено, что при крейсерских скоростях оба вида используют как активную, так и пассивную жаберную вентиляцию, а при наиболее форсированном движении — только пассивную. Показано, что это дает значительный (до 30%) энергетический выигрыш при критических скоростях (Fredman, 1979). Во всех описанных случаях вода проходит сквозь жабры водном направлении; движений типа «вдох — выдох» у водных позвоночных нет. Соотношение дыхательных движений ротовой полости и жаберной крышки таково, что вода прокачивается через жабры практически непрерывно (рис. 11): давление в ротовой полости на протяжении почти всего дыхательного цикла выше, чем в жаберной.
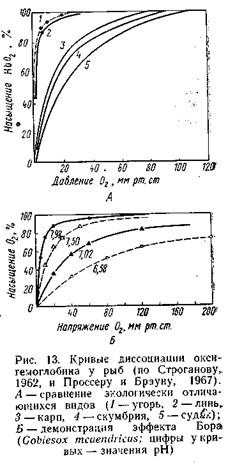
идет непрерывно, и отходящая от жабр кровь имеет почти тот же уровень насыщения О2, что и поступающая в жабры вода. Математический анализ принципов газообмена показал, что именно про тивоточная система обеспечивает наибольшую эффективность утилизации O2 (Piiper, Sheid, 1972). Благодаря всем рассмотренным особенностям дыхания некоторые костистые рыбы могут извлекать до 85% О2, растворенного в омывающей жабры воде. Велика у костистых рыб и степень утилизации О2, поступившего в кровь: она в 2,5—3 раза выше, чем у млекопитающих. У хрящевых рыб эффективность извлечения О2 из воды составляет 70—77% (при более быстром протоке 40—50%), что вполне сопоставимо с эффективностью извлечения О2 у костных рыб. От метим для сравнения, что у миног с их малоэффективным типом жаберного аппарата эта величина составляет 10—25%. Адаптации к колебаниям концентрации О2 в дыхательной среде осуществляются и на физиологическом уровне. В эксперименте за регистрировано, что на снижение количества О2 рыбы реагируют компенсаторным увеличением частоты дыхательных движений и (или) увеличением объема воды, пропускаемой через жаберный аппарат. При гипероксии дыхание, напротив, замедляется. Такая реакция имеет довольно общий характер: она отмечена у разных видов хрящевых и костных рыб, равно как и у круглоротых. Существенные адаптации к колебаниям напряжения О2 обнаруживаются в транспортной функции крови. Способность крови к переносу О2 обеспечивается, во-первых, растворением его в плаз ме, во-вторых — обратимым связыванием О2 с дыхательным пигментом крови (у позвоночных это гемоглобин). Количество О2, которое может быть растворено в плазме, ограничено; этот путь транспорта у большинства рыб имеет подчиненное значение. Известно лишь небольшое число форм, преимущественно антарктических (сем. Chaenichthydae), кровь которых полностью лишена гемоглобина, и весь О2 транспортируется только в плазме. Исследования, проведенные на одном из представителей этого семейства — ледяной рыбе Chaenocephalusaceratus, показали, что этот вид обитает в хорошо аэрированных холодных водах, ведет довольно пассивный образ жизни и обладает низким уровнем обмена веществ. Для него характерны обильное снабжение сосудами (васкуляризация) жабр, повышенная роль кожного газообмена, расширение сердца и увеличенная скорость кровотока в жаберных сосудах. Однако даже при этих свойствах устойчивость к гипоксии у ледяной рыбы (и у других рыб, лишенных гемоглобина) существенно ниже, чем у видов, имеющих гемоглобин. Кривая диссоциации оксигемоглобина (рис. 13), показывающая степень насыщения гемоглобина кислородом при различном его парциальном давлении, у наземных позвоночных обычно имеет сигмоидную форму, а у рыб — чаще гиперболическую, что свидетельствует о большем сродстве к кислороду. На этой кривой в качестве стандартных параметров приняты две точки: напряжение кислорода (в миллиметрах ртутного столба), при котором гемо глобин насыщается на 100% («зарядное напряжение»), и напряжение, при котором оксигемоглобин отдает 50% кислорода («разрядное напряжение», или «напряжение полунасыщения»).
При повышении сродства гемоглобина к кислороду кривая сдвигается влево (насыщение происходит при меньшем РО2). Этот эффект может зависеть от условий, в которых происходит газообмен; в частности, связи гемоглобина с кислородом ослабевают при повышении температуры. Большое значение имеет также величина рН: при повышении кислотности сродство гемоглобина к кислороду уменьшается и кривая диссоциации сдвигается вправо («эффект Бора»). Эффект Бора имеет очень большое значение при тканевом газообмене: имен но в тканях в кровь поступает большое количество СО2, рН понижается и гемоглобин начинает легче отдавать О2 тканям. Свойства гемоглобина у разных видов рыб могут довольно широко варьировать, отражая приспособления к кислородному режиму водоемов, в которых они обитают. В частности, у рыб, живущих в стоячих, богатых органикой водоемах, сродство гемоглобина к кислороду, как правило, выше, чем у обитателей быстротекущих вод или у неглубоководных морских рыб, которые не лимитированы количеством растворенного в воде кислорода (табл. 9).
Гематологические отличия связаны и с подвижностью: обитатели хорошо аэрированных водоемов обычно высокоподвижны; при этом пониженное сродство гемоглобина к кислороду, при котором он быстрее отдается в ткани, биологически оправдано. Рыбы из водоемов с низким содержанием кислорода, напротив, обычно ведут малоподвижный образ жизни, при котором быстрота отдачи . кислорода в ткани не столь существенна. Кроме того, у таких видов обычно хорошо выражен эффект Бора, облегчающий процесс тканевого газообмена. Экологические корреляции дыхательных свойств крови с естественными условиями кислородного режима прослеживаются и в пределах одного вида, в частности, в зависимости от возрастных особенностей. Показано, что личинки миног Lampetrafluviatilisотличаются более высоким сродством гемоглобина к кислороду (Р50 = 0,25 кПа), чем взрослые формы (Р50=1,43 кПа), что связано с обитанием личинок в грунте, т. е. в условиях затрудненного доступа кислорода (Bird et al, 1976). В реактивных ответах на гипоксию большую роль играет число эритроцитов: при появлении признаков кислородной недостаточности происходит выброс депонированных эритроцитов в кровь, что увеличивает ее кислородную емкость. Отмечено, что при этом рыбы, в естественных условиях испытывающие недостаток кисло рода, резче реагируют на возникновение гипоксии (табл. 10).
|