| Газообмен у ныряющих животных |
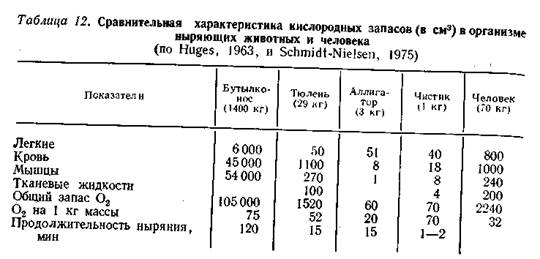
Своеобразные условия газообмена складываются у наземных животных, вторично перешедших к водному образу жизни или эко логически связанных с регулярным нырянием либо (как многие птицы) погружением головы в воду. Все эти виды имеют легочный тип дыхания, у всех процесс внешнего газообмена происходит в нормальных атмосферных условиях. Однако во время пребывания под водой резко изменяются условия снабжения тканей кислородом: поступление его в организм прекращается и возникает состояние функциональной асфиксической гипоксии. У наиболее специализированных ныряльщиков остановка дыхания может быть до вольно длительной, хотя у подавляющего большинства видов продолжительность пребывания под водой не превышает нескольких минут. Среди рептилий наибольшая (более часа) длительность нахождения под водой регистрируется у водных черепах и морских змей. Так, тихоокеанская зеленая черепаха Cheloniamtjdasвыдерживает погружение до 5 ч, а у бородавчатых змей Acrochordusgranulatusвремя естественного погружения доходит почти до 2 ч. У других водных рептилий продолжительность ныряния обычно не превышает 20-25 мин. Надо отметить, что у водных рептилий значительная часть газообмена при нырянии совершается водным путем через кожу и слизистые оболочки ротовой и анальной полостей (Хозацкий, 1967; Graham, 1974). Птицы по длительности погружения в большинстве случаев уступают водным млекопитающим. Длительность естественного погружения у них чаще всего меньше минуты. Ныряние буревестников, некоторых утиных, оляпки длится примерно до 15 с, у других утиных, а также у поганок - до 30 с. При насильственном погружении в воду (хотя бы только головы) тупик способен задержать дыхание на 4 мин, а чистики и кайры - даже до 12 мин. Пингвины (Pygoscelispapua, P. adeliae, Eudipteschrisolophus) при есте ственном нырянии находятся под водой 5-7 мин, а императорский пингвин Aptenodytesforsteri- до 18 мин. Ныряние вызывает довольно быстрое уменьшение содержания О2 и нарастание концентрации СО2 в крови. Устойчивость ныряющих животных к такой функциональной гипоксии определяется рядом физиологических адаптации, направленных, с одной стороны, на увеличение запасов О2 в организме перед нырянием, а с другой- на более экономное расходование этих запасов во время погружения. В результате относительная роль О2, содержащегося в воздухе, заполняющем легкие, в общем его запасании на время погружения оказывается второстепенной (табл. 12). У обыкновенного тюленя из общего запаса О2 в организме (40 мл/кг) только 14 мл/кг приходится на долю легочного запаса, тогда как у человека и собаки соотношение обратное: только 10 мл/кг приходится на внелегочные запасы.
Утки и, видимо, большинство других птиц также ныряют на выдохе. Только для пингвинов (Pygoscelisadeliae, P. papua) доказано ныряние на вдохе; при кратковременности погружения, свойственной этим видам, такая ситуация не грозит им опасным насыщением крови инертными газами. В то же время экспериментально показано, что запас воздуха в дыхательной системе обеспечивает возможность этим птицам продолжать газообмен в легких во время ныряния (Kooyman et al., 1973). Существенно большее значение имеет для ныряющих животных запасание О2 в крови (табл. 12). Эффективность этого пути определяется общим количеством крови, содержанием гемоглобина, числом и суммарной поверхностью эритроцитов и некоторыми другими гематологическими параметрами, в совокупности определяющими кислородную емкость крови. Отмечено, что эти параметры у хорошо ныряющих животных (китообразные, ластоногие) несколько выше, чем у неныряющих. Это прослеживается и в ряду ныряющих животных: исследование трех видов дельфинов показало, что общее содержание О2 в крови высокоактивной и глубоко ныряющей пелагической белокрылой морской свиньи Phocaenoidesdalliпочти в 3 раза выше, чем у прибрежной афалины Tursiopstruncatus, и на 70% больше, чем у пелагического полосатого дельфина Lagenorhynchusobliquidens, отличающегося не очень высокой активностью. Сходная картина обнаруживается и у птиц: параметры красной крови у ныряющих форм обычно несколько выше, чем у неныряющих. Так, общий объем крови у хохлатых пингвинов в среднем составляет 14,3% от массы тела, у хохлатой чернети-14,6, у красноголового нырка- 15,6, у гаги- 16-17%, тогда как у других водоплавающих около 13%, а у наземных птиц-в среднем около 6% от массы тела. Концентрация гемоглобина у пингвинов колеблется в пределах 17-20 г%, у тупика и кайры этот показатель составляет соответственно 18 и 19,4 г%, а у наземных видов он существенно меньше: у неясыти 7,7%, у домового воробья 10,8, у степного орла 12,6 г% (Коржуев, 1964; Коржуев и др., 1962, 1977). В результате у птиц-ныряльщиков несколько выше может быть и общая кислородная емкость крови: у пингвинов порядка 20 об. %, у чистиковых 24-26, у домашней утки около 17, а у кур 11,2 об.%; в то же время у голубя кислородная емкость крови составляет 21,2 об.%, что связано с активным полетом (Andersen, 1966). По признаку сродства гемоглобина к О2 ныряющие животные в большинстве случаев существенно не отличаются от неныряющих. Поскольку дыхание у всех этих животных происходит при нормаль ном давлении О2, повышение степени сродства не имело бы смысла; при разрядке оксигемоглобина в тканях высокое сродство к О2 скорее, затрудняло бы этот процесс. Относительно высокое сродство гемоглобина к О2 обнаружено у пингвинов, хотя и у них вели чина разрядного напряжения pso (28,8-34,4 мм рт. ст., или 3,8- 4,6 кПа) сопоставима с тем же показателем столь хорошего летуна, как голубь (29,5 мм, или 3,9 кПа). Не исключено, что высокое сродство гемоглобина к О2 выполняет у пингвинов особую функцию, способствуя ускорению реоксигенации крови при коротких появлениях кормящихся птиц на поверхности воды, а относительно выраженный эффект Бора облегчает снабжение тканей О2. Как уже говорилось, миоглобин обладает сродством к О2 при мерно в 10 раз большим, чем гемоглобин. Поэтому при обычных условиях он легко насыщается О2, транспортируемым с кровью, а в период погружения, когда приток О2 с кровью сокращается или прекращается совсем, отдает связанный ранее О2 ткани, окислительная ферментная система которой способна насыщаться О2 при малых величинах его парциального давления (рис. 19). Приспособления к экономному расходованию запасов кислорода. Как показывают расчеты, общий запас О2 в легких, крови и. мышцах не в состоянии обеспечить продолжительную остановку дыхания, свойственную ныряющим животным, если он расходуется с такой же скоростью, что и при свободном дыхании. Так, у пингвинов запас О2 перед погружением обеспечивает существование на уровне метаболизма покоя в течение 3 мин, тогда как длительность естественной задержки дыхания при нырянии доходит до 5-7 мин. Тюлень находится под водой втрое дольше, чем «позволяют» расчетные данные; примерно таково же соотношение расчетных и реальных величин у китов. Все это свидетельствует о том, что расход О2 в организме во время ныряния существенно ниже, чем при нормаль ном дыхании. Система приспособлений, определяющих этот эффект, имеет очень важное значение для всех ныряющих животных. В основе этих приспособлений лежат сердечно-сосудистые реакции организма.
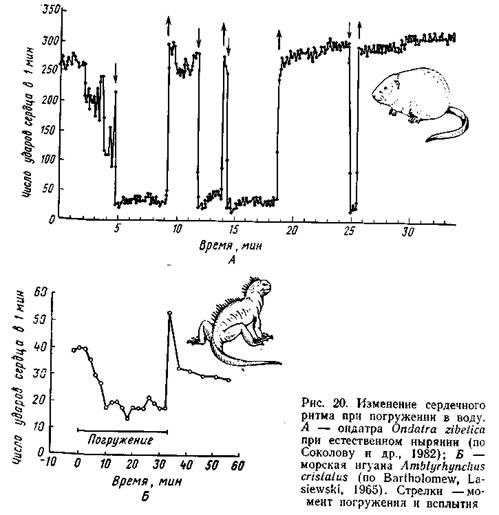
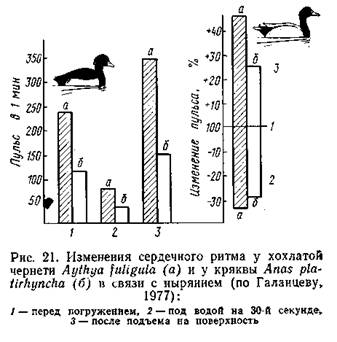
В опытах с тюленями и морскими львами, обученными нырять с укрепленными на теле электродами, было установлено, что при произвольном нырянии частота пульса сначала быстро, а затем замедленно снижалась до 20 ударов в минуту; у тех же животных при насильственном погружении пульс очень быстро (за 2-3 с) падал .до 8 ударов в минуту (Eisner, 1970). Аналогичные данные получены на птицах. В опыте с домашними утками насильственное погружение в воду вызывало урежение сердечного ритма до 1/4-1/20 от исходного уровня. Телеметрические наблюдения за свободноныряющими красноголовыми нырками и хохлатыми чернетями да ли несколько иные показатели: перед погружением отмечались тахикардия (учащение пульса) и учащенное дыхание, при погружении развивалась брадикардия, после чего частота пульса по степенно восстанавливалась до исходной величины, предшествовавшей всплытию (Butler, Woakes, 1979). Степень замедления сердечного ритма у разных видов неодинакова. У дельфинов при кратковременных произвольных заныриваниях частота сокращений сердца снижалась вдвое, у тюленя Уэддела (насильственное погружение) - в 3 раза, у морского слона (опыты с погружением головы) - в 4-6 раз, у каланов погружение в воду замедляет сердечный ритм в 2-8 раз. В результате замедления сердечной деятельности обмен не сколько снижается. Так, обыкновенный тюлень, в спокойном со стоянии расходующий около 200 мл О2 в минуту, при нырянии затрачивает всего 50 мл/мин. Показано, что частота пульса меняется Даже у одной особи в зависимости от характера ныряния: у тюле ней Уэддела глубокая брадикардия выражена лишь при длительных погружениях, а при кратковременных ныряниях частота пульса меняется слабо (Kooyman, 1975). Чрезвычайно существенно приспособление к экономному расходу О2 в виде изменения характера циркуляции крови в организме. Еще в 1940 г. П. Шоландер (Scholander, 1940) обнаружил, что в течение длительного ныряния содержание молочной кислоты в крови тюленей, уток и пингвинов повышается всего в 2-3 раза, но в момент всплытия концентрация ее в 10 раз превышает исходный (до ныряния) уровень. На этом основании он высказал предположение, что образующаяся в мышцах при гликолизе молочная кис лота не поступает в кровь из-за ограничения кровоснабжения мышц во время ныряния; при всплытии циркуляция в мышцах восстанавливается, и молочная кислота в большом количестве переходит в кровяное русло. Последующие исследования подтвердили эту гипотезу. В частности, с помощью ангиографии было показано, что на время ныряния у тюленей периферические кровеносные сосуды сжимаются и кровоток в них прекращается (или сильно замедляется). При этом основная масса мускулатуры практически отключается от снабжения крови кислородом. Особенно показательны исследования К. Иогансена (Johansen, 1964), который погруженным в воду уткам внутривенно вводил радиоактивный изотоп 86Rb в момент появления брадикардии. Через 2 мин уток забивали и исследовали распределение изотопа в тканях, что служило надежным показателем уровня кровотока в разных частях организма. В результате было установлено, что во время ныряния резко уменьшается (вплоть до полного прекращения) снабжение кровью большей части мускулатуры (кроме мышц головы и пищевода, которые у утки деятельно функционируют при сборе пищи под водой), кишечника, кожи (кроме кожи в области головы), почек, поджелудочной железы. Мозг и сердечная мышца, судя по этим опытам, снабжаются кровью нормально, а в щитовидной железе и надпочечниках отмечено несколько повышенное кровоснабжение. Практически все данные по сердечно-сосудистым адаптациям к нырянию получены в опытах с насильственным погружением в воду испытуемого животного или только его головы. Немногие исследования на пингвинах (Адели, антарктический) с использованием радиотелеметрической техники позволили получить сведения о реакциях организма при естественном нырянии. Было выяснено, что брадикардия выражена и при естественном погружении пингвинов: частота сердечных сокращений в погруженном состоянии составляла у пингвина Адели 25%, а у антарктического - 70% от исходного (до заныривания) уровня. В отличие от уток у пингвинов отмечено снабжение кровью мускулатуры, хотя и сниженное (Millard et al., 1973). Как указывалось, пингвины ныряют на вдохе, и, по-видимому, во время ныряния продолжается газообмен запасенного воздуха с кровью. У совершающих короткие (около 1,5 мин) заныривания дельфинов афалин, приученных к условиям опыта в открытом море, так же обнаружена внутримышечная циркуляция крови (Ridaway, Howard, 1979). Вероятно, этот тип адаптации используется лишь при длительном погружении. Принципы сердечно-сосудистых адаптации в виде возникновения брадикардии и перераспределения тока крови оказываются общи ми для ныряющих животных. Перераспределение кровотока во время ныряния обеспечивает поддержание нормального, несмотря на брадикардию, артериального давления, а также подачу О2, запасенного в крови, только к органам, активно работающим в погружен ном состоянии. Остальные органы (в первую очередь мускулатура) используют запасы О2 в виде оксимиоглобина, а отчасти переключаются на анаэробные процессы, о чем свидетельствует подчас значительное повышение уровня молочной кислоты как в самих мышцах, так и (после всплытия) в крови. Биологическая значимость перераспределения кровотока при нырянии подтверждается тем, что в опытах с тюленями выключение сфинктера задней полой вены вело к гибели подопытных животных уже после 4-минутного пребывания под водой (Packer et al., 1969). Сложная система адаптивных реакций ныряющих животных контролируется центральной нервной системой на уровне продолговатого мозга. В экспериментах на птицах двусторонняя перерезка тройничного нерва снимала рефлекс брадикардии и остановки дыхания при обычных для этих реакций «пусковых механизмах», о которых говорилось выше. Аналогичный эффект вызывала перерезка или холодовой блок блуждающих нервов (Andersen, 1963; Butler, Jones, 1968). Более детальных сведений о механизмах нервной регуляции сердечно-сосудистых реакций при нырянии, так же как о природе рецепторов, не имеется. |