Водно-солевой обмен у водных животных |
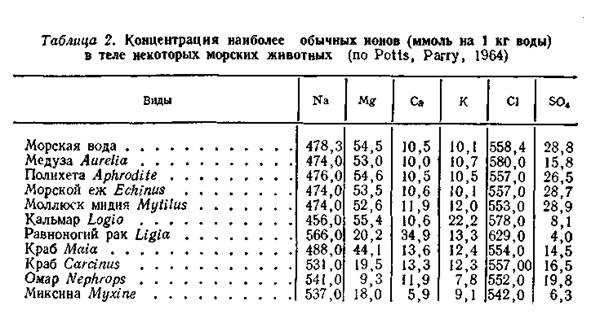
У первичноводных животных жаберный, кишечный, а иногда и покровный эпителий всегда контактирует с водой, поэтому через их покровы постоянно осмотическим путем проникает вода, если концентрация солей (и других веществ) в организме отличается от таковой окружающей водной среды. Жизнь формировалась в морской среде, что наложило отпечаток на основные физико-химические особенности животного организма. В частности, ионный состав жидкостей тела качественно сходен с составом морской воды даже у наземных животных. Однако количественное содержание различных ионов в теле даже морских животных может отличаться от соотношения их в окружающей среде (табл. 2).
Объясняется это способностью организмов избирательно извлекать из среды и задерживать в своем теле определенные соли. Некоторые из них идут на построение скелета, раковин и иных «инертных» образований, другие циркулируют в составе жидкостей тела. Ионы некоторых солей, напротив, активно выводятся из организма. Следует различать ионную регуляцию в организме, связанную с поддержанием состава и количественного соотношения различных ионов в жидкостях тела, и регуляцию осмотического давления, которое не связано с этими показателями, а определяется суммой растворенных частиц. Ионная регуляция свойственна практически всем животным, осмотическая же регуляция может отсутствовать. В этом случае осмотическое давление внутри организма следует за изменениями давления внешней среды; таких животных называют пойкилоосмотичными (осмоконформерами). Животные, способные к активной регуляции осмотического давления, поддерживают относительное постоянство этого параметра внутренней среды независимо от окружающей воды; таких животных называют гомойоосмотическими (осморегуляторами). Первичноводные морские беспозвоночные относятся к первой группе. Осмотическое давление жидкостей их тела практически равно таковому морской воды, изменяясь вместе с изменением ее солености. Такие животные называются изотоничными. Абсолютная изотоничность свойственна относительно немногим группам (кишечнополостные, иглокожие); у большинства же беспозвоночных отмечается некоторое превышение осмотического давления жидкостей тела (гипертоничность организма), что обеспечивает постоянный приток воды в организм в пределах, легко уравновешивающихся процессами выделения. Способность изотоничных животных переносить некоторые изменения солености среды определяется главным образом механизмами клеточной устойчивости к обводнению и дегидратации. Диапазон устойчивости таких организмов обычно не очень велик, поэтому изоосмотические осмоконформеры распространены, как правило,. в морских водоемах с относительно устойчивой соленостью. Беспозвоночные-осморегуляторы могут жить в условиях большего диапазона солености среды. Это обеспечивается механизмами активной регуляции осмотического давления внутренней среды. Эти механизмы включают изменения проницаемости мембран, активный перенос ионов, а также изменения внутриклеточной концентрации свободных аминокислот в направлении, уравновешивающем суммарное осмотическое давление в клетке с внешней средой.
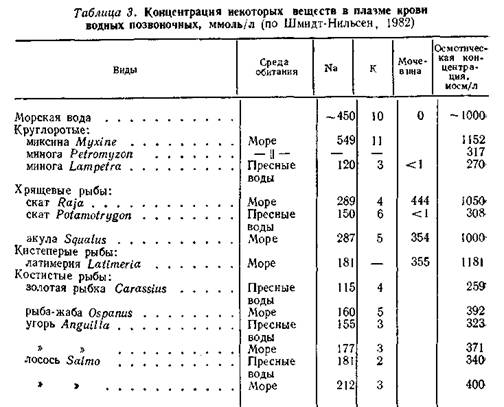
Пресноводная осморегуляция. В организме пресноводных животных концентрация жидкостей внутренней среды обычно несколько ниже, чем у морских форм, но все же выше, чем в окружающей воде (Концентрация солей в пресных водоемах колеблется в пределах 0- 0,5
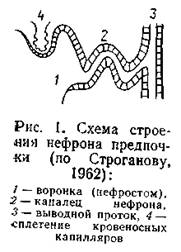
Благодаря этому вода осмотическим путем постоянно поступает внутрь организма. У ряда пресноводных животных различные покровные образования затрудняют проникновение воды через кожу (панцири, чешуя и т. п.). Однако полная изоляция организма от осмотического поступления воды невозможна, поскольку по меньшей мере эпителий ,• органов дыхания и слизистой кишечника неизбежно контактирует с водой. Это вызывает необходимость специальных адаптации, направленных на выведение из организма избытка воды. В основном такие адаптации связаны с функцией выделительной системы. Хорошим примером развития подобных приспособлений могут быть первичноводные позвоночные, в частности крутлоротые и рыбы. Предки их были морскими животными, но собственно позвоночные (подтип Vertebrata) эволюционно формировались как обитатели пресных вод. Наиболее эффективная адаптация к обитанию их в гипотонической среде - образование почки. Почка водных позвоночных с самого начала формировалась не только как орган выделения, но и как орган осморегуляции (тем более, что продукты белкового обмена этих животных - аммиак и мочевина - растворимы в воде и выводятся главным образом внепочечным путем, через жабры). В индивидуальном развитии первичноводных позвоночных сменяется два типа структуры почки. Вначале развивается так называемая головная почка (pronephros). Она состоит из большого числа канальцев, каждый из которых открывается в полость тела воронкой, снабженной ресничками. Внешне эти канальцы (нефроны) похожи на нефридии кольчатых червей или бесчерепных, но расположены не метамерно, а собраны в единый целостный орган и своим вторым концом открываются в общий выводящий проток (рис. 1).
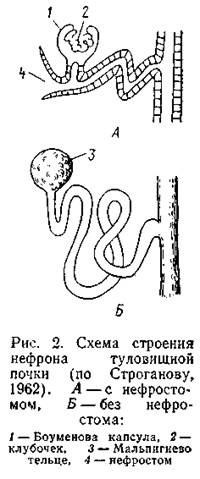
Движения ресничек вызывают ток жидкости из полости тела внутрь канальца и далее, через общий проток, наружу. Канальцы предпочки выстланы железистым эпителием и несут экскреторную функцию. В ряде случаев вблизи воронок в стенках брюшной полости образуются сплетения кровеносных капилляров, сквозь стенки которых вода и некоторые растворенные в ней вещества проникают в полость тела, попадая отсюда в воронки нефронов. Предпочка функционирует как зародышевый орган (Есть предположение, что предпочка функционирует у взрослых миксин, но вопрос этот до конца не ясен), у взрослых форм развивается новая серия нефронов, образующих туловищную почку (mesonephros). Внутреннее строение туловищной почки сходно с предпочкой и отличается в первую очередь тем, что в начальной части нефронов, вблизи воронки, стенки канальца образуют выпячивание (боуменова капсула), внутри которого заключен клубочек кровеносных капилляров (гломерулус); это образование называется мальпигиевым тельцем (рис. 2, А).
У рыб часть нефронов утратила воронки, и они прямо заканчиваются мальпигиевыми тельцами, не имея связи с полостью тела (рис. 2, Б). Канальцы нефронов этого типа обычно более длинные, извитые; некоторые их участки приобрели функцию всасывания. В клубочке силой кровяного давления жидкая часть плазмы крови вытесняется из капилляров в просвет боуменовой капсулы, откуда попадает в извитой каналец нефрона. В состав этой жидкости (первичная моча) входит не только вода, но и растворенные в ней вещества, частицы которых могут проникнуть сквозь стенки капилляра (ионы солей, молекулы Сахаров, мочевина, мочевая кислота и др.); практически лишь белки, молекула которых слишком крупна, не попадают в состав первичной мочи. Процесс образования первичной мочи нередко называют ультрафильтрацией. В извитом канальце нефрона, который оплетен сетью кровеносных капилляров, происходит обратное всасывание солей, сахара и ряда других веществ в кровяное русло. В результате конечная моча гипотонична по отношению к плазме крови. В целом клубочковая система почки функционирует по фильтрационно-реабсорбционному типу, выделяя гипотоническую мочу, что вполне соответствует принципам осморегуляции в пресной воде. Клубочковая система - главная морфологическая основа интенсивной фильтрации и выведения воды из организма. Воронки с их ресничками выводят воду со значительно меньшей скоростью. Благодаря клубочкам пресноводные рыбы выводят очень большое количество слабоконцентрированной мочи.
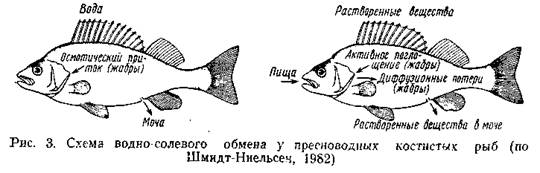
Выведение избытка воды еще не гарантирует постоянства осмотического давления внутренней среды. Организм непрерывно теряет часть солей в составе мочи и экскрементов, а также в результате диффузии через кожу. Восстановление необходимых солей происходит с пищей. Кроме того, важную роль в солевом обмене играет жаберный эпителий, специализированные клетки которого способны к активной абсорбции солей (главным образом одновалентных ионов Na+ и С1-) из окружающей воды. В целом принцип осморегуляции пресноводных рыб сводится к следующему (рис. 3): вода поступает в организм осмотическим путем через жабры и слизистую пищеварительного тракта; избыток ее выводится через почки. Активного питья воды не происходит.
Если поступают с пищей и через жабры; потеря их идет с мочой, экскрементами и частично через кожу. Фильтрационно-реабсорбционная функция почек, так же как и абсорбционная активность жаберного эпителия, зависят от соотношения осмотических давлений воды и жидкостей организма. Это позволяет пресноводным рыбам осваивать водоемы с некоторой степенью осолонения, и таким образом экологические возможности отдельных видов расширяются. Аналогично осуществляется пресноводная осморегуляция и у беспозвоночных животных: у них также основная функция выведения воды принадлежит выделительной системе; при этом изменение солености внешней среды адекватно изменяет уровень выделительной функции.
Неизбежные потери солей с мочой и экскрементами и в этом случае компенсируются захватом ионов из окружающей воды. У некоторых животных этот процесс идет через всю поверхность тела, но главную роль играют жабры и аналогичные образования. Показано, в частности, что так называемые «анальные жабры» личинок ряда видов насекомых в первую очередь служат органом осморегуляции, возможно вообще не участвуя в дыхании.
Таким образом, в отличие от морских беспозвоночных все пресноводные животные гомойоосмотичны; активные процессы осморегуляции способствуют поддержанию относительного постоянства солености внутренней среды. Поэтому большинство пресноводных форм может обитать и в солоноватоводных водоемах (соленость 0,5-30
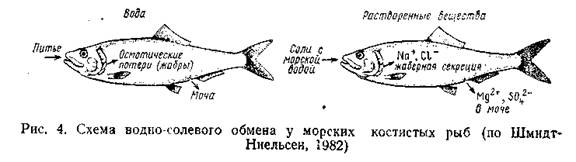
Осморегуляция в море; костные рыбы. Освоение позвоночными морской среды началось, по-видимому, в конце силура - начале девона, когда в результате морской трансгрессии многие материковые водоемы оказались связанными с океаном и осолонились. Условия водно-солевого обмена в морской и пресной воде кардинальным образом отличаются: в море организм животных оказывается гнпотоничным по отношению к среде (соленость океанических вод составляет 30-40%, в среднем - 35 Приспособления к осморегуляции в таких условиях развивались в разных группах позвоночных на основе пресноводного принципа строения почки, свойственного их предкам. При этом у большинства морских костных рыб функциональная активность клубочковой системы снижается; значительная часть клубочков вообще не участвует в фильтрации. В опытах с форелью Saltno gairdneri было показано, что хотя фильтрационная активность отдельных нефронов в морской воде выше (3,74 нл/мин против 1,31 нл/мин в пресной воде), общая гломерулярная фильтрация за счет уменьшения числа функционирующих нефронов в морской воде ниже, чем в пресной (соответственно 20,1 и 142,6 нл/мин). У многих морских костистых рыб уменьшается размер клубочков, а нередко и их число. Наконец, среди морских костистых рыб есть и виды, совершенно утратившие клубочки: канальцы нефронов в таких агломерулярных почках заканчиваются слепо и обладают активной секреторной функцией. Агломерулярная почка характерна, в частности, для рыб семейства Syngnathidae, а также встречается у ряда антарктических видов. В последнем случае утрата клубочков, по-видимому, способствует сохранению в крови гликопротеинов, играющих важную роль «биологических антифризов» (см. гл. 3). Снижение уровня почечной фильтрации не в состоянии полностью скомпенсировать осмотические потери воды. Поэтому морские рыбы регулярно пьют воду, получая при этом избыточное количество солей. Экспериментально выяснено, что угорь и морской подкаменщик, находящиеся в морской воде, поглощают до 50- 200 см3 воды. Если блокировать возможность ее поступления через кищечник, рыба обезвоживается и после потери 12-20% исходной массы погибает; введение таким рыбам воды через анальное •отверстие снимает эффект обезвоживания. В опытах с лососем Salmo gairdneri рыбы, содержавшиеся в пресной воде, не пили, а в 32, 50 и 100%-ной морской воде поглощали соответственно 42, 95 и 129 мл воды на 1 кг в сутки; 80% выпитой воды абсорбировалось в кишечнике. Реабсорбция ионов в почечных канальцах морских рыб снижена, зато в отличие от пресноводных здесь происходит обратное всасывание воды из канальцевой мочи. Соответственно морские рыбы выделяют сравнительно небольшое количество мочи, концентрация которой почти равна (лишь немного ниже) плазме крови. Интенсивно всасывается вода и в мочевом пузыре костистых рыб, стенки которого обладают высокой осмотической проницаемостью. Кроме того, как и у пресноводных рыб, здесь абсорбируется хлористый натрий. Избыток солей, помимо почек, выводится через кишечник: здесь происходит интенсивное всасывание воды, в то время как соли концентрируются и выводятся с фекалиями. Это относится главным образом к двухвалентным ионам, тогда как хлористый натрий активно абсорбируется, создавая в слизистой оболочке градиент концентрации, вследствие которого вода переносится через стенку кишечника. Большую роль в выведении избытка солей играют жабры. Если двухвалентные ионы в значительном количестве выводятся через почки и пищеварительный тракт, то одновалентные (главным образом Na+ и Сl-) экскретируются почти исключительно через жабры, выполняющие у рыб двойную функцию - дыхания и выделения. В жаберном эпителии есть особые крупные бокаловидные клетки, содержащие большое число митохондрий и хорошо развитый эндоплазм этический ретикулум. Эти «хлоридные» клетки расположены в первичных жаберных лепестках и в отличие от дыхательных клеток связаны с сосудами венозной системы. Перенос ионов через жаберный эпителий имеет характер активного транспорта и идет с затратой энергии. Стимулом экскреторной деятельности хлоридных клеток является повышение осмолярности крови. Участие жабр в активном транспорте одновалентных ионов характерно не только для костистых рыб; хлоридные клетки обнаружены у миног, акуловых рыб, хрящевых и костных ганоидов. Как указывалось, активный перенос ионов свойствен и пресноводным рыбам, но происходит у них в обратном направлении. Не до конца еще ясно, функционируют ли в этом случае другие клетки, аналогичные хлоридсекретирующим, или же меняется «знак» работы последних; по наиболее современным данным вероятнее второе предположение. В целом схему осморегуляции морских костистых рыб можно вставить следующим образом (рис. 4).
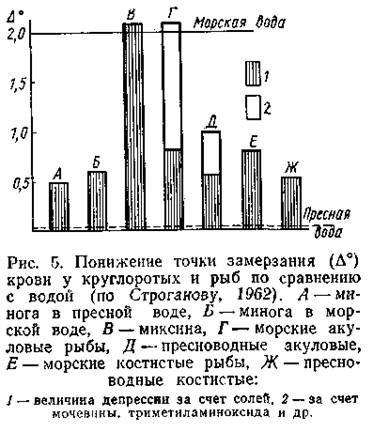
Гипертоничность внешней среды обуславливает постоянные осмотические потери воды Основном через жабры), которые восполняются путем питья. Из-за получаемых при этом солей выводится через почки и с фелиями (главным образом двухвалентные ионы), а также активно экскретируется особыми клетками жаберного эпителия (в основном Na+ и Сl-). Функционирование механизмов осморегуляции позволяет поддерживать осмотическое давление внутренней среды на относительно постоянном уровне, гипотоническом по отношению к внешней среде. Осморегуляция у хрящевых рыб. Хрящевые рыбы приспособились к обитанию в морской среде совершенно иным путем. Концентрация солей в их крови близка к таковой у костистых рыб и ниже, чем концентрация их в морской воде (рис. 5).
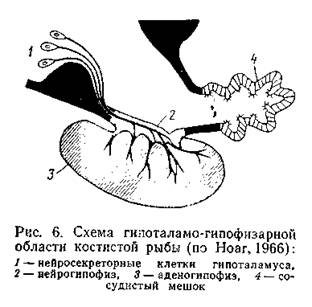
Однако осмотическое давление жидкостей тела у этих рыб практически равно осмотическому давлению морской воды, лишь слегка превышая его (табл. 3). Достигается это тем, что в почечных канальцах хрящевых рыб активно реабсорбируется мочевина, 70-99% которой возвращается в кровь, повышая ее суммарное осмотическое давление. Проницаемость жабр для мочевины у этих рыб в отличие от костистых понижена, и избыток мочевины выводится практически только через почки. Помимо мочевины, в крови хрящевых рыб накапливается значительное количество триметиламиноксида (ТМАО), также обладающего высокой осмотической активностью. ТМАО содержится в жидкостях тела многих морских организмов, но у акуловых рыб его концентрация особенно велика. Так, если у морских костистых рыб содержание ТМАО в крови составляет 25-460 мг%, то у акуловых оно достигает 250-1430 мг%. До сих пор не ясно, продуцируется ли это вещество в организме или поступает в него с пищей. На снижение солености среды хрящевые рыбы реагируют Уменьшением реабсорбции мочевины и усилением выведения ее (и ТМАО) с мочой. Благодаря этим регуляторным процессам акуловые рыбы (по крайней мере, некоторые виды) выдерживают большие колебания солености среды, появляясь даже в пресных водах. Относительно немногие пресноводные скаты, имея клубочковую почку, осуществляют осморегуляцию, подобную костистым пресноводным рыбам. Хотя содержание мочевины в крови у них выше, чем у костистых рыб, все же оно значительно меньше, чем у морских форм; реабсорбция мочевины в почечных канальцах практически отсутствует, жабры способны поглощать ионы натрия и хлора из окружающей среды. Изоосмотичность морских пластиножаберных рыб не означает отсутствия ионной регуляции. Напротив, содержание солей в составе внутренней жидкой среды поддерживается на довольно постоянном уровне. Количество хлористого натрия в крови акул и скатов почти в два раза ниже, чем в морской воде, несмотря на постоянный приток этих ионов с пищей и благодаря диффузии через жабры. Часть избыточных солей выводится с мочой: концентрация ионов калия, кальция, магния и некоторых других в проксимальных сегментах нефронов нарастает (Stolte et al., 1977). Хлористый натрий, напротив, реабсорбируется в дистальных почечных сегментах нефронов и возвращается в кровь. Некоторое количество этих ионов выводится хлоридными клетками жабр, но главную роль в экскреции хлористого натрия, видимо, играет ректальная железа- пальцеобразный вырост, соединенный протоком с клоакой. Показано, что до 75% выводимого из организма хлористого натрия экскретируется ректальной железой. При искусственной солевой нагрузке секреторная активность этой железы повышается; при помещении подопытных акул в опресненную воду концентрация электролитов в секрете ректальной железы уменьшается. Поскольку внутренняя среда организма" хрящевых рыб слегка гипертонична по сравнению с морской водой, происходит умеренный осмотический приток воды в организм, который обеспечивает потребности мочеобразования и секреции ректальной железы; акуловые рыбы в отличие от костистых не пьют морскую воду. Аналогичный хрящевым рыбам тип осморегуляции обнаружен у единственного современного вида кистеперых рыб Latimeria chalumnae, ведущего морской образ жизни. Благодаря высокому (сравнимому с акулами) содержанию в крови мочевины (355 ммоль/л) и ТМАО (более 100 ммоль/л) общая осмотическая концентрация плазмы латимерии близка к таковой вод Индийского океана (соответственно 1181 и 1000 мосм/л). При этом концентрация электролитов составляет лишь 40% от их концентрации в морской воде. Сходство с хрящевыми рыбами дополняется наличием у латимерии ректальной железы (Pickford, Grant, 1967; Lutz Robertson, 1971). Значительное количество мочевины синтезируется и задерживается в крови у двоякодышащих рыб. При этом отмечается определенная экологическая зависимость: у лепидосирена, переживающего неблагоприятный период во влажной среде, уровень синтеза мочевины занимает промежуточное положение между протоптерусом (находится в сухом коконе) и неоцератодусом, постоянно плавающем в воде (Funkhouser et al., 1972). У протоптеруса обнаружена лоакальная железа, хотя и несходная с ректальными железами якул и латимерии, но, видимо, играющая активную экскреторную поль в период пребывания в коконе (Lagios, McCosker. 1977). Р у осетровых рыб в морской воде также увеличивается содержание осмотически активных веществ сыворотки крови, что ведет выравниванию осмотического давления крови и окружающей среды. В частности, при адаптации байкальского осетра и большого амударьинского лопатоноса к воде с соленостью 10,5°/оо содержание мочевины в крови возрастало более чем в 2 раза. При этом в отличие от акуловых рыб у ряда осетровых колеблется и содержание солей, в частности хлористого натрия. Создается впечатление, что осморегуляция у осетровых рыб происходит по типу, промежуточному между осморегуляцией хрящевых и костистых рыб. Среди крутлоротых миксины - типичные морские формы - характеризуются тем, что жидкости их тела изотоничны морской воде. У этих животных более 99% осмотического давления внутренней среды определяется неорганическими ионами, концентрация которых быстро выравнивается с таковой окружающей среды (чаще слегка превышая его, как у акуловых рыб). Нередко это обстоятельство рассматривают как показатель пойкилоосмотичности миксин. Исследования, однако, показывают, что миксины Eptatretus stouti способны к активной реабсорбции натрия в почечных канальцах (Mclnerney, 1974). Об активной осморегуляции говорит и то, что искусственная осмотическая стимуляция гиперосмотическим раствором вызывает у миксин Myxine glutinosa активацию гипоталамо-гипофизарной системы (Vogel et al., 1971). Миноги обладают активной осморегуляцией. Как пресноводные, так и морские виды имеют клубочковую почку; концентрация солей в их теле ниже, чем в морской воде (см. рис. 5). Механизмы осморегуляции миног в принципе такие же, как у костистых рыб. Хотя большинство морских беспозвоночных изотоничны, есть и формы, которые, подобно рыбам, гипотоничны и осуществляют в морской среде активную осморегуляция. Таковы, в частности, некоторые ракообразные пресноводного происхождения. Замечателен в этом отношении рачок Artemia salina, обитающий в соленых озерах. Как и морские рыбы, артемии заглатывают соленую воду, активно выделяя избыток солей через жабры. Благодаря этому, хотя жидкость в пищеварительном тракте рачка всегда содержит много солей, гемолимфа сохраняет гипотетичность по отношению к среде. Физиологический контроль осморегуляции. Активная осморегуляция обеспечивает не только приспособления принципиального характера (регуляция в пресной или морской воде), но и лабильные адаптивные реакции на изменение градиента солености между организмом и внешней средой. Это в значительной мере расширяет экологические возможности активных осморегуляторов. Интенсивность функций осморегуляторных механизмов стимулируется динамикой осмотического давления внутренней среды; у рыб регуляция этих процессов связана главным образом с нервно-гуморальными механизмами системы гипоталамус - гипофиз--интерреналовая ткань (Интерреналовая ткань - гомолог коры надпочечников высших позвоночных, которые у рыб не выражены. Эта ткань состоит из железистых клеток, развивающихся из целомического эпителия боковых пластинок). Гипоталамус представляет собой центральную структуру головного мозга, в которой сигналы с осморецепторов трансформируются в адаптивные реакции организма. Наиболее прямая форма адаптивного ответа заключается в том, что вырабатываемые в ответ на осмотическую стимуляцию нейросекреты гипоталамуса переносятся по аксонам в заднюю долю гипофиза (нейрогипофиз), откуда в виде гормонов поступают в кровь. У рыб в нейрогипофизе продуцируются гормоны аргинин-вазотоцин (АВТ), изотоцин (ИТ), окситоцин (ОТ) и вазопрессин (ВП). Первые три гормона в низких дозах (до 1 нг/кг) действуют в пресной воде как антидиуретики, а в морской воде их влияние на диурез не проявляется или проявляется в виде слабой стимуляции мочеотделения. В высоких же дозах (более 10 нг/кг) все эти гормоны повышают диурез. Вазопрессин до недавнего времени у низших позвоночных не обнаруживался и считался гормоном, специфичным для млекопитающих. Недавно его обнаружили у рыб, у которых этот гормон обладает диуретическим действием, причем в пресной воде он подавляет антидиуретический эффект АВТ, ИТ и ОТ (Следует иметь в виду, что одни и те же гормоны в разных группах животных действуют неодинаково. В частности, вазопрессин у млекопитающих выступает как актидиуретический гормон. Еще более разнообразно действие гормона пролактина (см. ниже)). Все нейрогипофизарные гормоны влияют на диурез, воздействуя на интенсивность клубочковой фильтрации. У круглоротых (миноги), по-видимому, вырабатывается только вазотоцин; он, возможно, секретируется в очень малых количествах. Другая форма влияния гипоталамуса на водно-солевой обмен заключается в стимуляции эндокринной деятельности передней доли гипофиза (аденогипофиз). Проникающие в аденогипофиз (рис. 6) нейросекреты стимулируют определенные группы клеток, продуцирующих гормоны. Наибольшее значение в осморегуляции имеют пролактин и адренокортикотропный гормон (АКТГ).
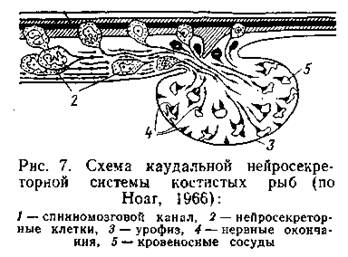
Пролактин играет ведущую роль в пресноводной осморегуляции. Поэтому он секретируется у пресноводных рыб в большем количестве, чем у морских; при переводе морских рыб в среду с пониженной соленостью продукция пролактина существенно возрастает. В экспериментах с удалением гипофиза инъекции пролактина обеспечивают подопытным рыбам возможность нормальной осморегуляции в пресной воде. В морской воде пролактин определяет уменьшение размера почечных клубочков (в пресной - увеличение х), а также выступает как антагонист кортизола (см. ниже) в регуляции абсорбции воды и ионов хлористого натрия в кишечном тракте. Стимуляция других клеток аденогипофиза влечет за собой увеличение секреции АКТГ, в свою очередь повышающего активность нтерреналовой ткани, клетки которой выделяют ряд кортикостероидов, принимающих непосредственное участие в регуляции водно-солевого обмена. В иитерреналовой ткани костистых рыб вырабатывается не менее шести кортикостероидов, а также ряд их про4зводных. Видимо, наиболее важное значение в регуляции водно-солевого обмена играет кортизол (У высших позвоночных кортизол, кортизон, кортикостерон в первую очередь регулируют обмен углеводов), в меньшей степени - альдостерон. Показано, что гипофизэктомированные угри при содержании морской воде не в состоянии повышать абсорбцию воды в кишечнике и более чем в 4 раза замедляют скорость обращения Na+ через жабры; аналогичные нарушения наблюдаются и при интереналэктомии. Инъекция кортизола восстанавливает осморегуляторные свойства. Вообще система АКТГ - кортизол считается лавным фактором приспособления рыб к осморегуляции в морской воде. Судя по некоторым работам, у акулообразных и осетровых рыб гормональный контроль осморегуляторных процессов осуществляется аналогично. Есть сведения, что определенную роль в регуляции водно-солевого обмена у рыб играют и гормоны, выделяемые клетками хромаффиновой ткани (Хромаффиновая ткань - гомолог мозгового слоя надпочечников высших поззвоночных - развивается из зачатков симпатической нервной системы), особенно адреналин (эпинефрин). Адреналин увеличивает проницаемость жаберного эпителия для воды; одновременно снижается выведение хлоридов и натрия через жабры. У акулы Squalus acanthias внутримышечное введение адреналина вызывало усиление диуреза и уменьшение содержания в крови мочевины и ТМАО (Forster et al., 1972). Помимо интерреналовой и хромаффиновой ткани, в почках лучеперых рыб обнаруживаются отдельные группы железистых клеток, заключенные в соединительнотканные капсулы - тельца Станниуса. Функция их не до конца ясна, но последние исследования показывают, что эти образования участвуют в регуляции обмена кальция. Помимо гипоталамо-гипофизарной, для рыб характерна еще каудальная нейросекреторная система: она находится в заднем от деле спинного мозга, где расположены нейросекреторные клетки, отростки которых оканчиваются в его брюшной части. Участок спинного мозга, в котором происходит накопление секрета и последующее выведение его в кровь, носит название урофиза (или нейрофиза). У костистых рыб урофиз вычленяется морфологически в виде вентрального выроста спинного мозга (рис. 7).
У хрящевых и осетровых рыб он морфологически не выражен, однако система нейросекреторных клеток имеется и у них. Структурная организация урофиза и нейрогипофиза сходны. Функции каудальной нейросекреторной системы, видимо, многообразны и изучены не полно. Есть сведения и об участии этой системы в регуляции водно-солевого обмена: введение экстрактов урофиза карпа лабораторным крысам продемонстрировало антидиуретический эффект (Ishikawa, 1980). Показано, что урофиз влияет на уровень ионов натрия, магния и хлора в плазме крови (Fryer et al., 1978).
Механизмы регуляции водно-солевого обмена у рыб находятся' в сложном взаимодействии, что определяет возможность лабильного изменения аеморегулятормых процессов в зависимости от динамики внешних условий и внутреннего состояния организма. Адаптивные возможности проявляются у разных видов неодинаково, как правило, в соответствии с особенностями среды их обитания и образа жизни. Так, рыбы Xiphister atropurpuretis, живущие в приливно-отливной зоне, систематически подвергаются колебаниям солености среды и соответственно быстро и совершенно регулируют свой водно-солевой обмен. Три вида бычков p. Cortus показывают различную способность к ионному и осмотическому обмену, что связано с их биологией; в экспериментах при повышении солености воды скорость потери натрия через жабры у пресноводного вида С. morio была примерно вдвое ниже, чем у морских С. bubalis и С. scorpius. В этих же условиях С. morio и С. bubalis (обитающий в опресненных участках моря) увеличивают объем выпитой воды, тогда как чисто морской вид С. scorpius, встречающийся в местах с устойчивой соленостью, не проявляет такой реакции (Foster, 1969). Аналогичным образом популяция Cyprinodon variegatus, живущая в открытой лагуне с резко меняющейся соленостью, оказалась более устойчивой к искусственно создаваемым перепадам концентрации, чем популяции того же вида, обитающие в условиях хотя и различной (16 и 290/00), но устойчивой солености (Martin, 1968).
Это же характерно для беспозвоночных животных. Например, офиуры Ophiothrix angulata из эстуария в Южной Каролине (США) менее устойчивы к снижению солености, чем представитетого же вида из эстуария во Флориде, что вполне соответствует разнице в условиях существования: в первом случае средняя соленость близка к морской (30%0) и колебания ее меньше, во втором наблюдаются частые и длительные периоды опреснения, и средняя соленость воды снижена примерно до 25 Замечательным примером широкой приспособляемости к солевому режиму водоемов служат так называемые проходные формы круглоротых (миноги) и рыб. При миграциях из моря в реки (или наоборот) механизмы осморегуляции у этих животных полярно-преобразуются (смена гипо- и гипертонического состояния организма), что основано на пресноводном типе структуры почек у всех водных позвоночных.
У молоди лососевых рыб процесс подготовки к смене типа осморегуляции начинается еще в реке: увеличивается концентрация осмотически активных веществ в плазме крови, возрастает число хлоридных клеток в жабрах и их ферментативная активность и т. д. Все это повышает устойчивость к возрастающей солености воды при выходе в море. У взрослых особей, напротив, при миграциях к устьям рек перестройки носят обратный характер, что позволяет им подняться вверх по течению на места нереста. Все эти изменения стимулируются нейроэндокринными механизмами, рассмотренными выше (Баранникова, 1975). В целом сложная система осморегулирующих механизмов определяет не только общую адаптацию водных животных к жизни в водоемах определенного типа, но и приспособления к использованию различных экологических ниш в каждом варианте водной среды, в том числе и возможность обитания в условиях неустойчивой (эстуарии крупных рек, приливно-отливные зоны и пр.) или резко отличающейся солености. и верхних горизонтов почвы. Все амфибии развиваются в водной среде, личинка их - типично водное животное. Связь амфибий с водой помимо прочих причин обусловливается типом водно-солевого обмена, который у этих животных основывается на тех же морфофункциональных механизмах, что и у пресноводных рыб. Туловищная почка амфибий выполняет главную функцию выведения излишка воды, легко проникающей осмотическим путем через голую кожу. Поступление воды через кожу с избытком обеспечивает потребности организма, поэтому амфибии практически ее не пьют или пьют очень мало (0,1-0,4 мл/кг-ч). При нахождении амфибий в воде в их почках усиливается клубочковая фильтрация, продуцируя значительное количество (10-25 мл/кг-ч) гипотонической мочи. Примерно половина фильтрующейся в клубочках воды может при необходимости реабсорбироваться; реабсорбция же ионов натрия и хлоридов практически полная (порядка 99%). При дегидратации реабсорбция воды достигает 94-95%. Кожные покровы амфибий способны к активному транспорту ионов из менее концентрированной водной среды внутрь организма - процесс, идущий с затратой энергии и направленный на компенсацию неизбежных потерь солей с мочой и экскретами. Поскольку большинство видов амфибий пресноводны, такой тип осморегуляции достаточно эффективен в отношении как водного, так и солевого баланса внутренней среды организма. Более того, как и у рыб, изменения уровня фильтрации и способность противостоять некоторому увеличению концентрации ионов в плазме крови обусловливает перенесение земноводными некоторого осолонения среды. При этом уменьшаются темпы клубочковой фильтрации, возрастает уровень канальцевой реабсорбции воды и снижается реабсорбция ионов натрия. В результате выделяется меньше мочи с повышенной концентрацией солей. Способность к осморегуляции у разных видов неодинакова. Среди европейских бесхвостых наибольшей устойчивостью к изменениям солености отличаются зеленая Bitfo viridis и камышевая В. calamita жабы. Опыты с американскими безлегочными саламандрами Batrachoseps relictus и В. atenuatus показали, что популяции этих животных, обитающие вблизи моря, более выносливы к повышению солености воды, чем популяции из удаленных от побережья районов. Способность к солевой адаптации обнаружена и в ряде экспериментальных исследований. Определенную роль в этом играют лимфатические мешки, которые заполняются большим количеством жидкости, изоосмотичной по отношению к плазме крови. Наиболее эвригалинные формы бесхвостых амфибий могут повышать концентрацию мочевины в крови - тип адаптации, сходный с таковым у хрящевых рыб. Так, у зеленой жабы при повышении осмотического давления среды от 55 до 806 мосм/л концентрация мочевины в крови увеличивалась в 6 раз. Совершенно уникальна в этом отношении лягушка-крабоед Rana cancrivora, обитающая в мангровых зарослях Юго-Восточной Азии и постоянно контактирующая с морской водой. Содержание мочевины в крови этой лягушки достигает 350-480 ммоль/л. Как и у хрящевых рыб, осмотическое давление плазмы крови у этого вида несколько вы-ще, чем в окружающей среде, что определяет осмотическое поступление воды в количестве, обеспечивающем необходимый уровень мочеобразования. Благодаря этому отпадает необходимость пить соленую воду (Gordon et al., 1961). В отличие от хрящевых рыб содержание мочевины в крови лягушки-крабоеда повышается не в результате канальцевой реабсорбции, а путем диффузии мочевины из мочевого пузыря. Интенсифицируется также синтез мочевины в организме, что доказывается активацией ферментов орнитинмочевинного цикла при повышении солености среды. Для этого вида установлено, что действие нейрогипофизарных гормонов направлено в основном на усиление проницаемости стенок мочевого пузыря для мочевины (Dicker, Elliott, 1973). При всем сходстве этого способа осморегуляции с осморегуляцией хрящевых рыб вряд ли его можно рассматривать как прямое «наследство» далеких предков. Показано, что у головастиков лягушки-крабоеда осморегуляция осуществляется по принципу, свойственному костистым рыбам (Gordon, Tucker, 1965). Оплодотворение яиц и процесс метаморфоза у этого вида не могут проходить в морской воде; эти критические моменты онтогенеза осуществляются только в периоды дождей, когда вода в мангровых болотах опресняется. Таким образом, оба способа осморегуляции выступают скорее как частная экологическая адаптация, открывающая виду своеобразную экологическую нишу. Амфибии - группа, занявшая наземные местообитания, но сохранившая при этом прежний тип водно-солевого обмена. В аналогичном положении и некоторые беспозвоночные животные, освоившие наземные наиболее влажные биотопы. Таковы, например, дождевые черви. Голые проницаемые покровы этих животных определили лимит пригодных местообитаний; они роются в почвенных горизонтах, насыщенных парами воды, которые абсорбируются покровами; прямой контакт их тела с капельно-жидкой влагой также служит постоянным источником увлажнения. Покровы дождевых червей легко проницаемы для воды в обоих направлениях; помещенные в сухом воздухе, черви теряют влагу и погибают. По характеру осморегуляции дождевые черви также близки к пресноводным животным. Если их поместить в раствор с меняющейся концентрацией, то у них устанавливается гипертоничность жидкостей тела по отношению к внешней среде и гипотоничность мочи по отношению к внутренней среде организма. Приспособления, ограничивающие дегидратацию
|